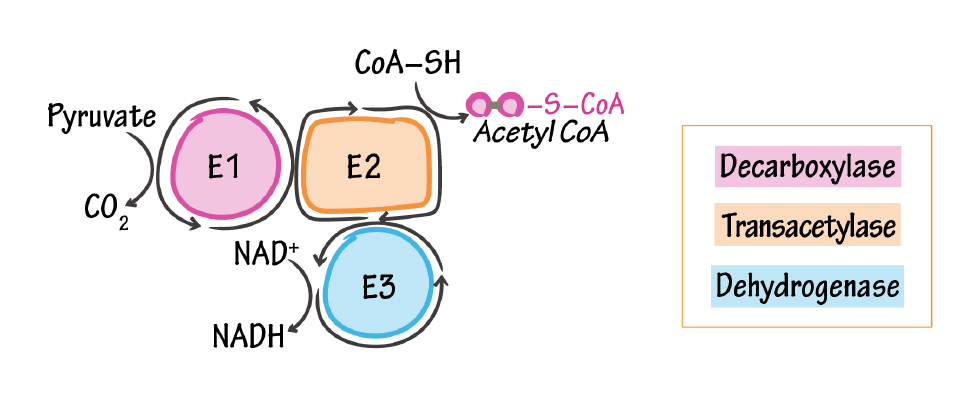
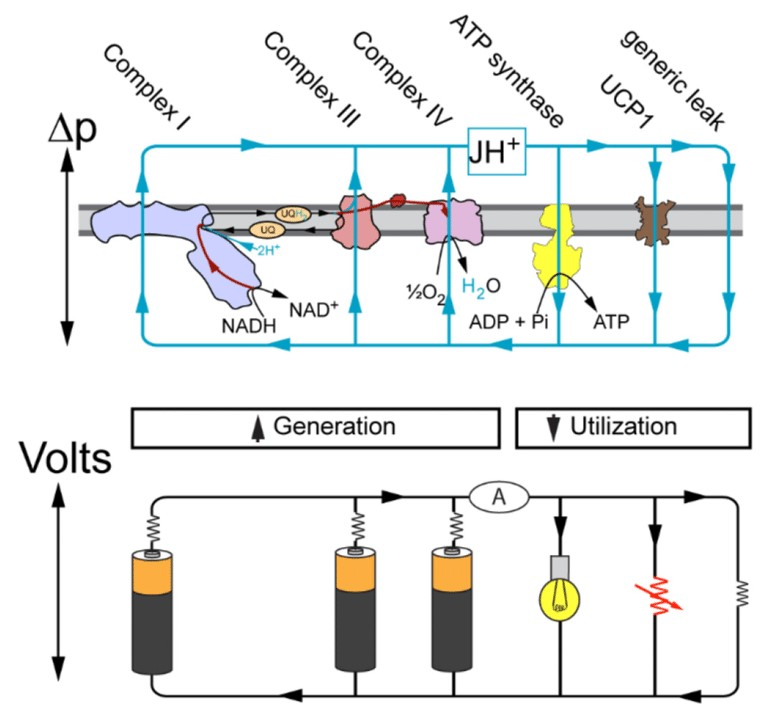
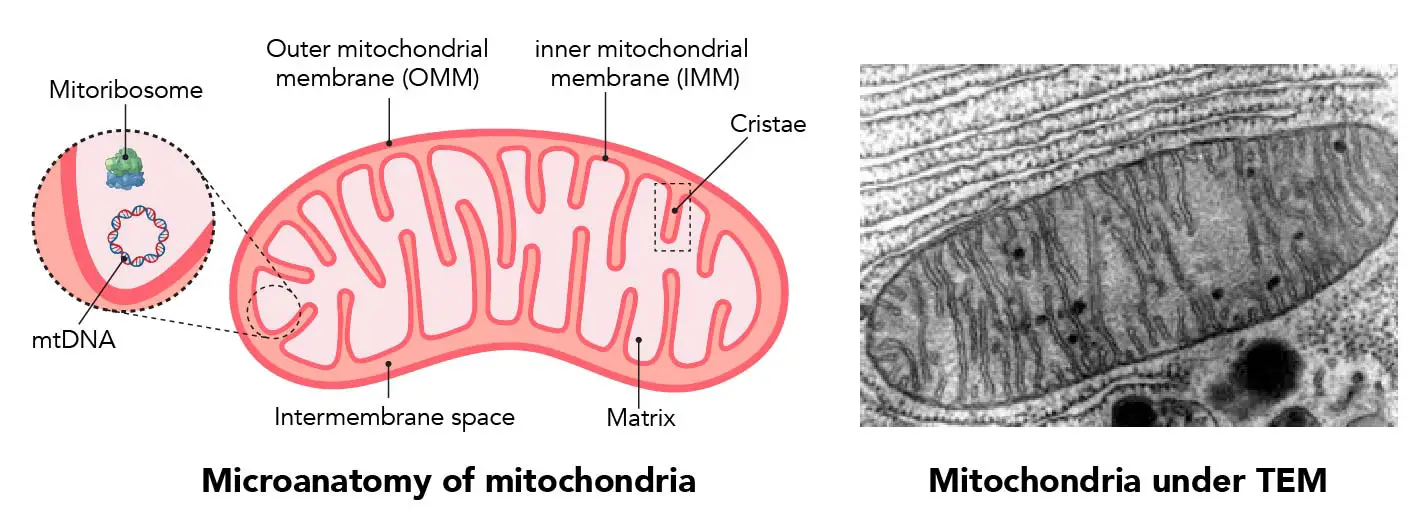
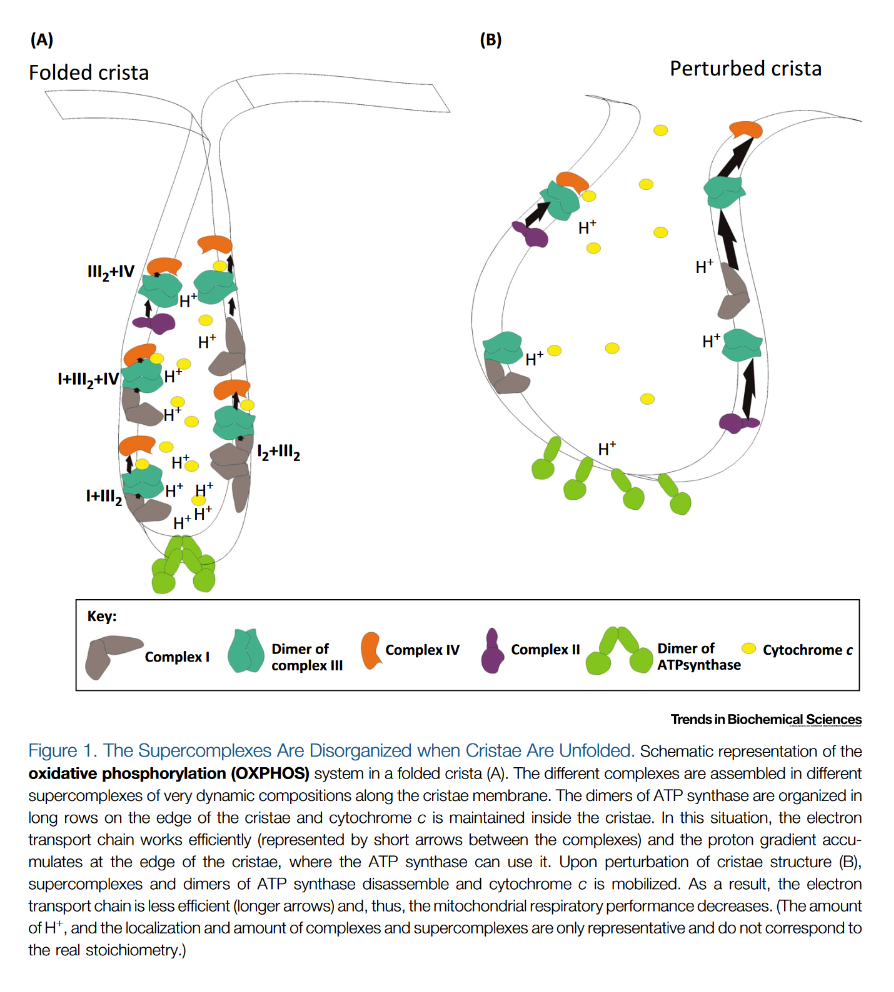
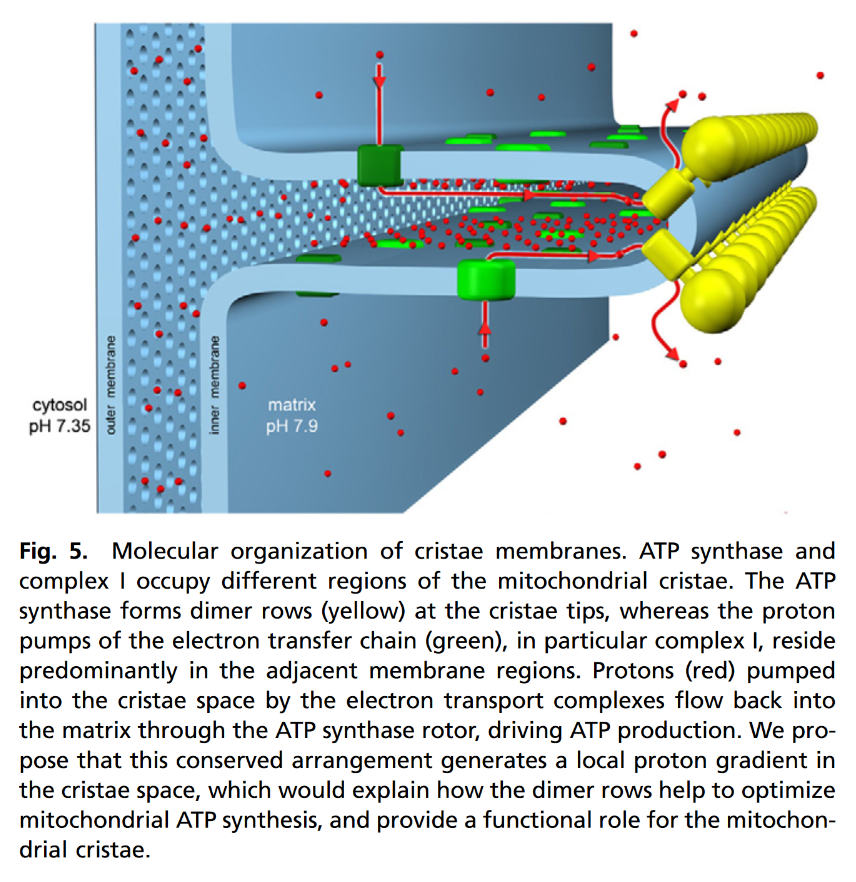
NADPH Oxidases (NOX): An Overview from Discovery, Molecular Mechanisms to Physiology and Pathology
This relates to the previous post and to the next information.
If you want to question your current understanding of cellular respiration, check out the work of Kelath Manoj and his subversive murburn model (stands for 'mild, unrestricted burning'), where he challenges anything that crosses the way.
- Murburn concept | Wikipedia
- Kelath Manoj | ResearchGate
- Kelath Manoj | Google Scholar
- Satyamjayatu (his foundation)
- Satyamjayatu | YouTube
⠀ - Murburn concept: A facile explanation for oxygen-centered cellular respiration
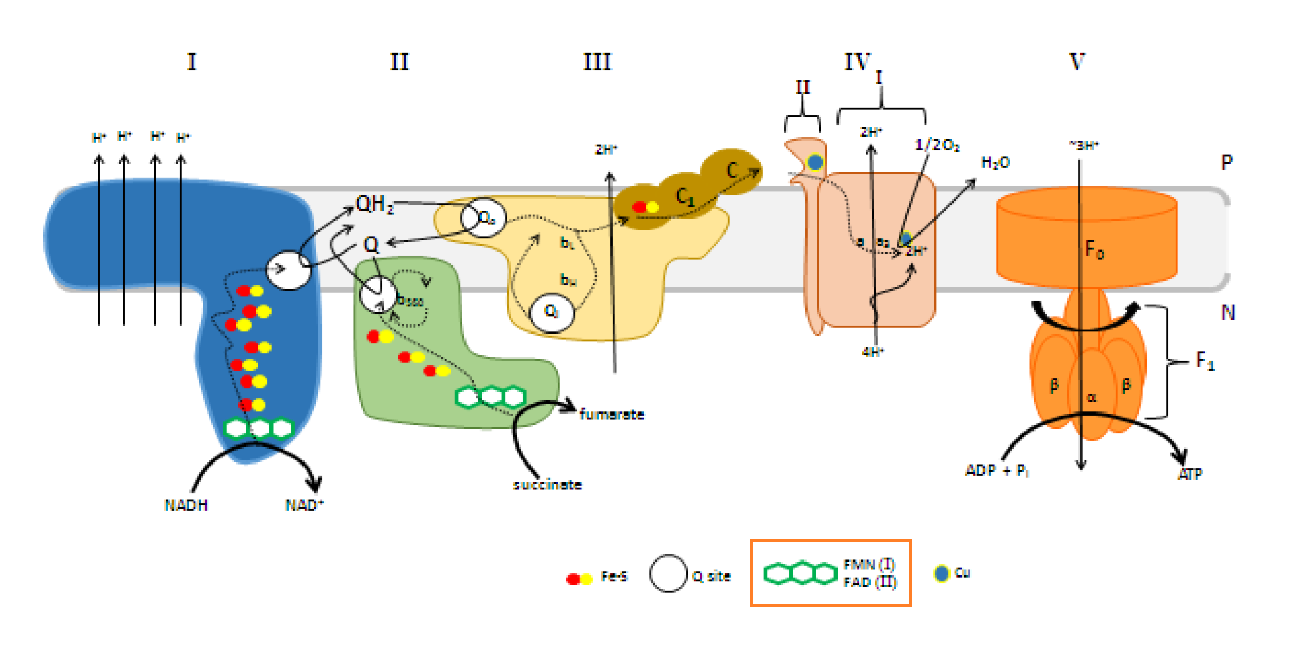
- Mitochondrial oxidative phosphorylation: Debunking the concepts of electron transport chain, proton pumps, chemiosmosis and rotary ATP synthesis
- Murburn concept and murzymes in 2023: Celebrating 25th year of pursuit
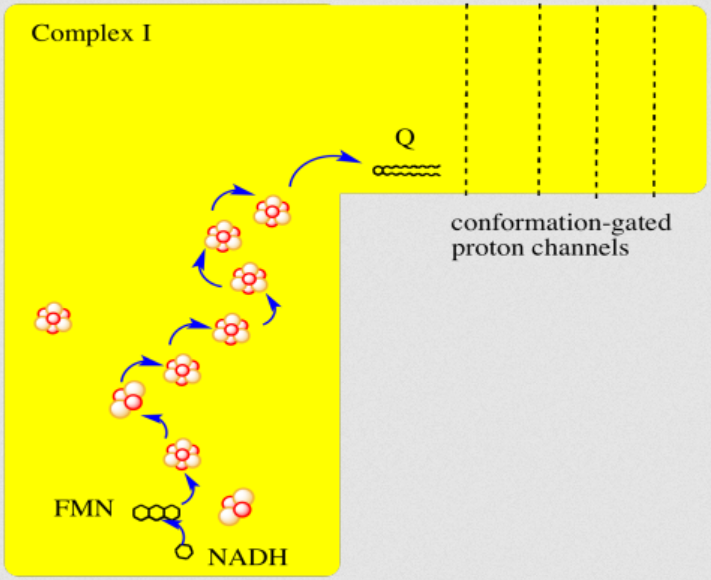
He treats (diffusible) reactive oxygen species (DROS) as an essential part of cellular respiration. Speculates that the projections of respiratory complexes were evolutionarily shaped to take advantage of radical generation. He identifies sites of ATP synthesis in all respiratory complexes and it's as oxidative phosphorylative as it gets: a reduction of oxygen to superoxide in respiratory complexes leads to its diffusion towards local ADP or phosphate attack, and a straightforward reaction joins them to form ATP without involving the convoluted steps of protons movement to drive ATP synthase (Complex V) subunit rotation.
- Superoxide radical as electron donor for oxidative phosphorylation of ADP (referenced by him)
Aerobic respiration: Proof of concept for the oxygen-centric murburn perspective
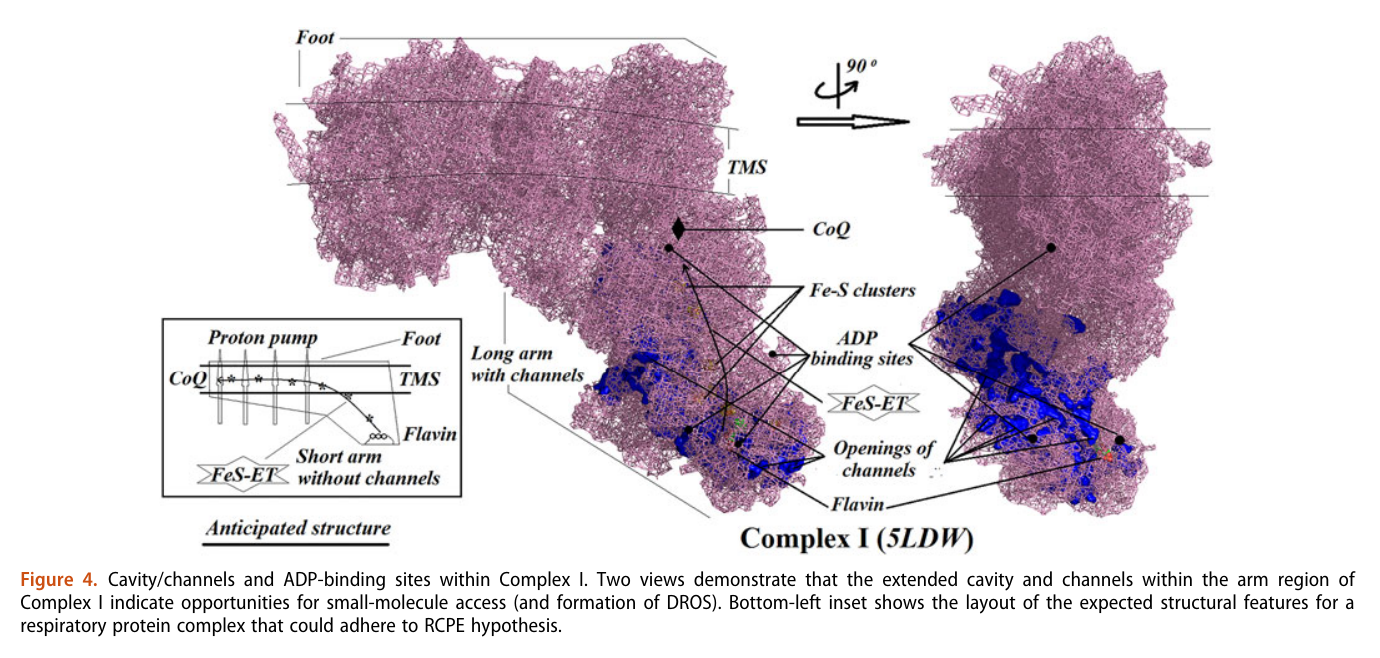
A respirawesome or supercomplex [I(III)2IV] showing 11 ADP-binding sites within projections into the matrix:
- 6 in Complex I
- 4 in Complex III
- 1 in Complex IV
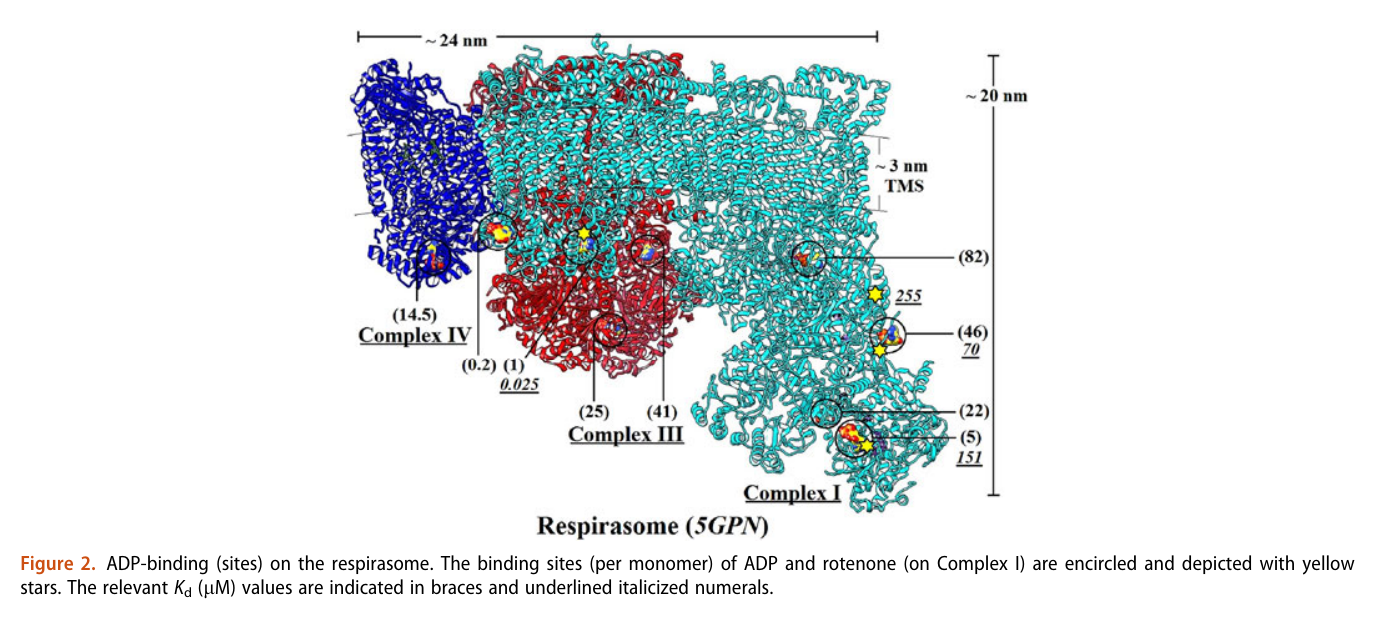
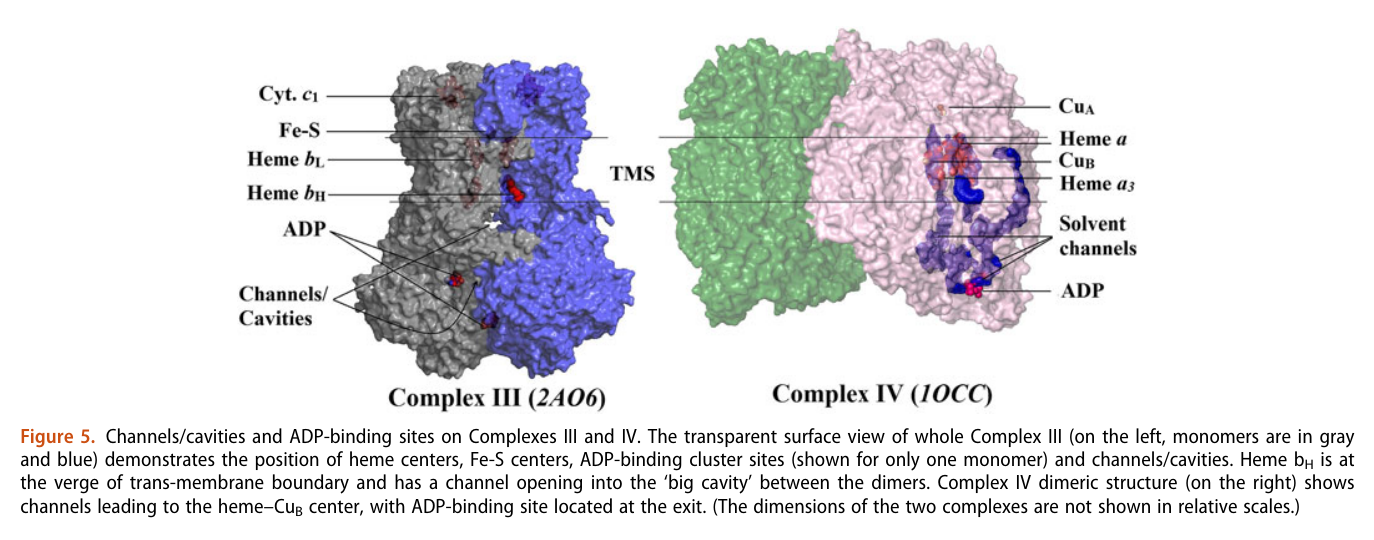
His team also report 'cavities or channels' that promote diffusion of oxygen species to desired targets (be them ADP, phosphorus or the known electron acceptors):
The proposed chain of reactions is shown here (omitting what occurs in between each):
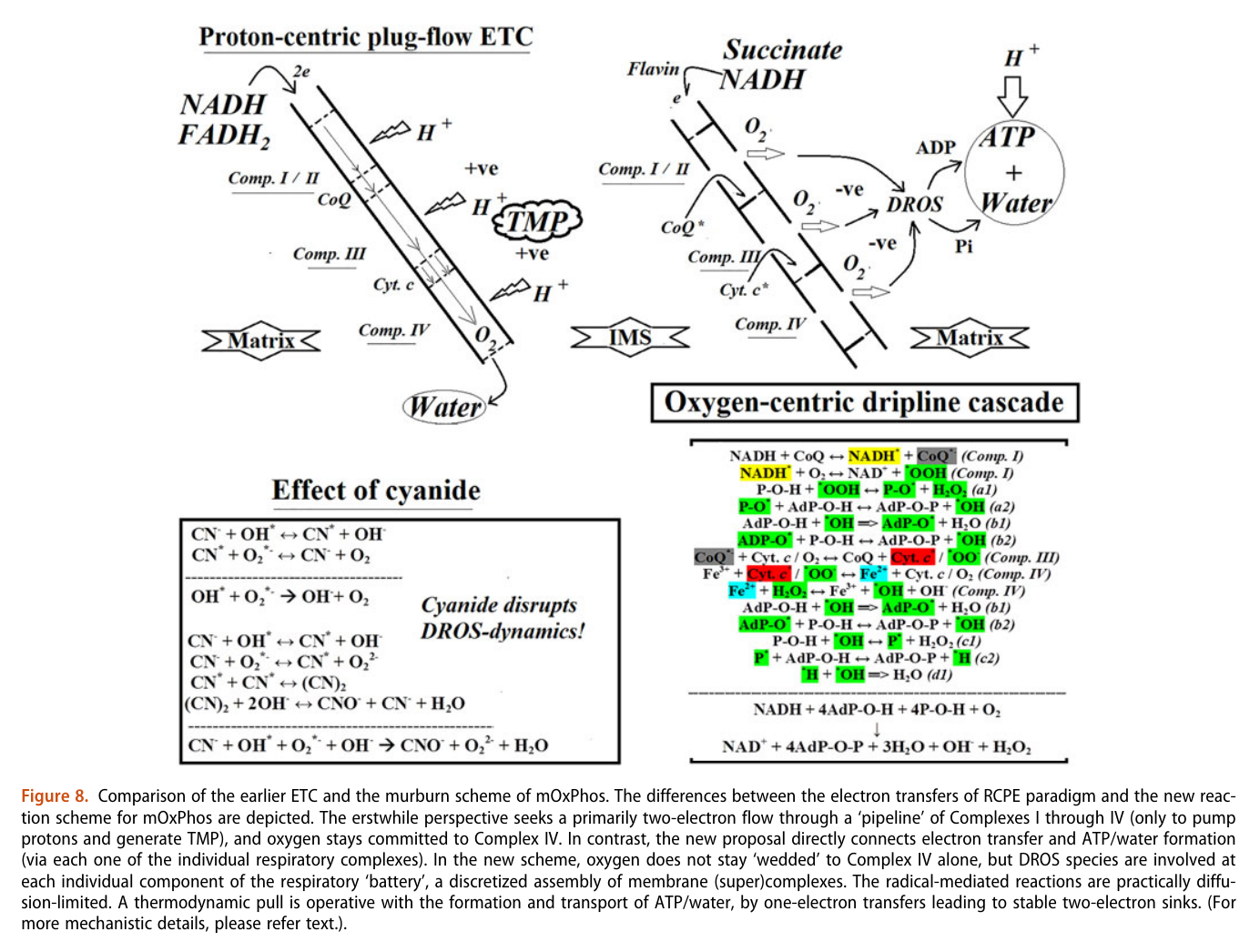
| AdP-O-H | ADP |
| P-O-H | Pi (inorganic phosphate) |
| AdP-O-P | ATP |
When complexes cluster, the organization favors their interaction, and ADP-binding sites from one complex can make use of diffusing reactive oxygen species from another. The strategic location and shared metabolites are shown in this diagram:
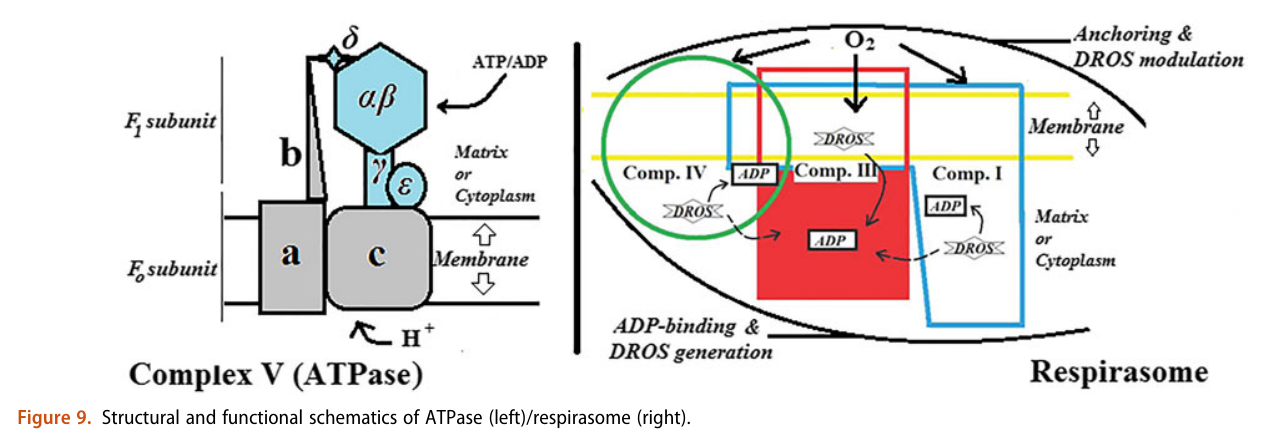
He not only diminishes the importance of Complex V as a source of ATP synthesis, but suggests that it's a decomposer of ATP (an ATPase). According to him, Complex V has much higher affinity for ATP than ADP, which is odd for a protein that's intended to produce ATP. To compound the issue, he argues that ADP is available at lower concentration than ATP.

The half-affinity for ADP being close to its concentration would be a problem.
He acknowledges the meek quantity of free protons per mitochondrion, which would be another concern for a 'rotary' protein dependent on them, but rejects the buffering or hopping possibility. I haven't looked into why. He adds to the picture the following considerations:
- Complexes per mitochondrion: ~10^4-10^5
- Proton requirement: ~10^6 (which is ×10 the previous value, assuming 10 H+ extruded per 1 NADH oxidized in the traditional model)
There's a lot more to uncover from his work. As an example, he's familiar with Gilbert Ling's model, compares it with his proposal and the traditional approach.