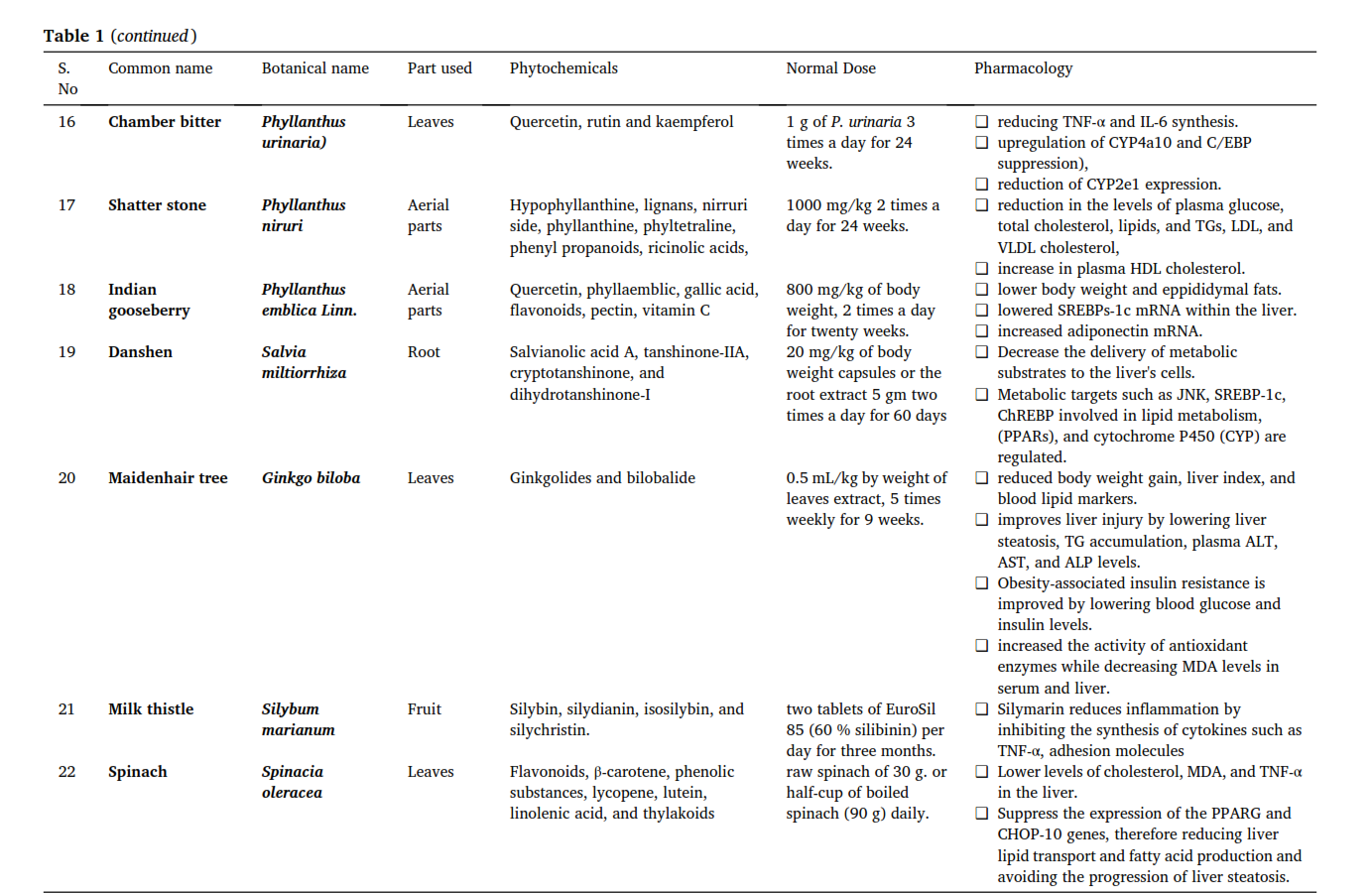
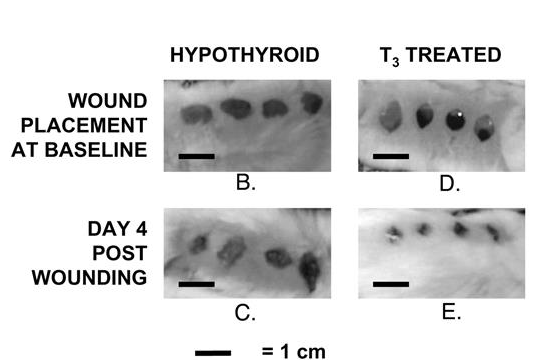
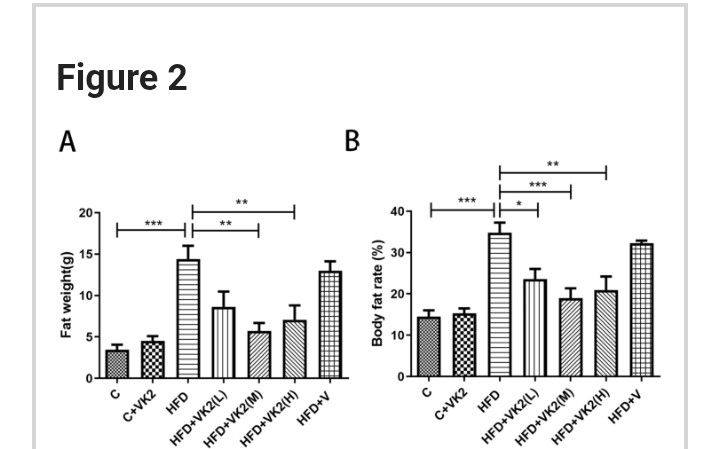
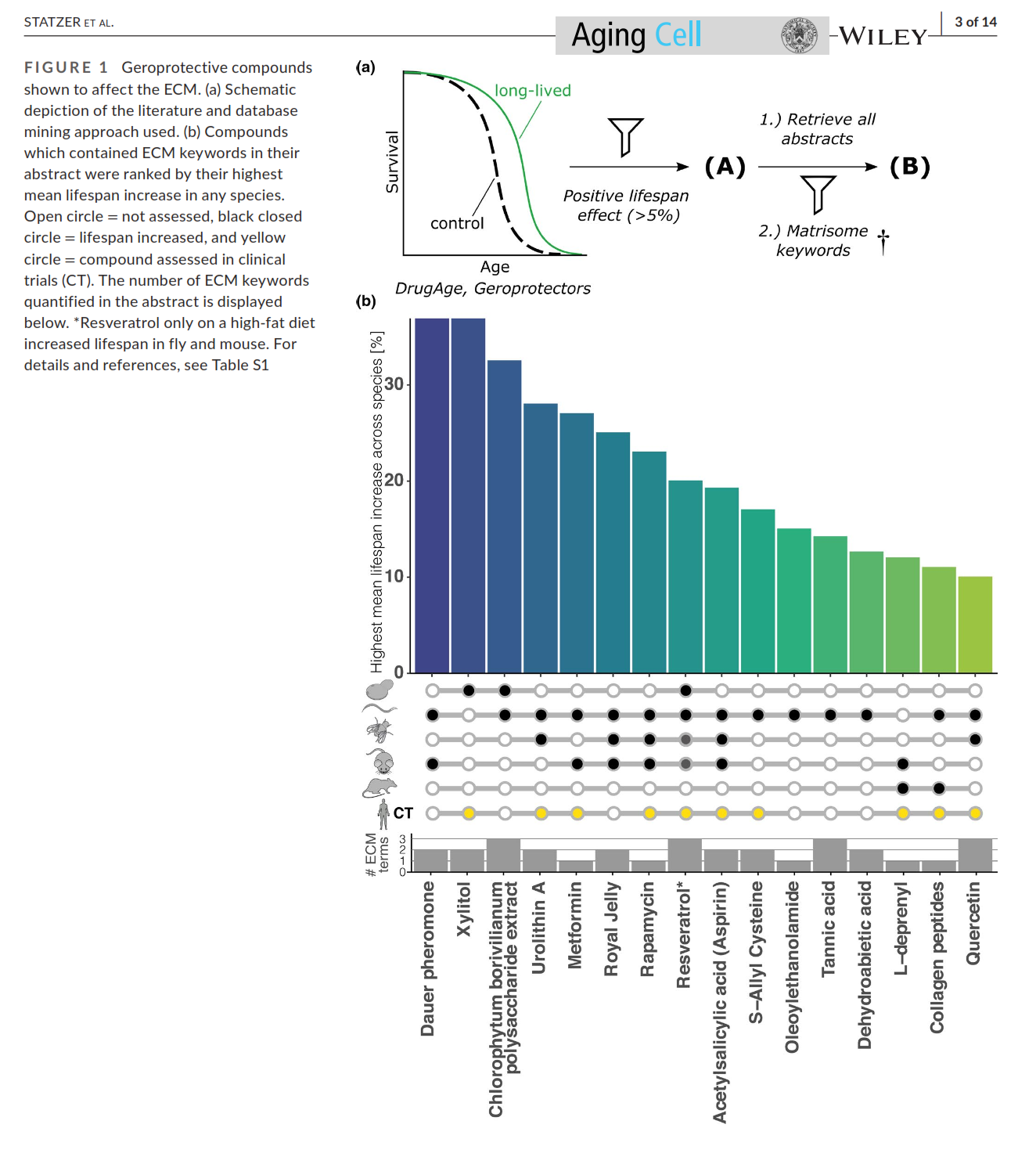
@Mossy said in Vitamin K2 reverses fatty liver disease:
do remember you mentioning this on another post, and I tried the calcium before MK4 — not to prevent palpitations, but just to have it not give me a bad reaction. It didn't work for me. Maybe it's needing to find the perfect ratio: calcium, magnesium, MK4, Vitamin D, Vitamin A, etc.
Thats odd, but thanks for the feedback.