@bio3nergetic
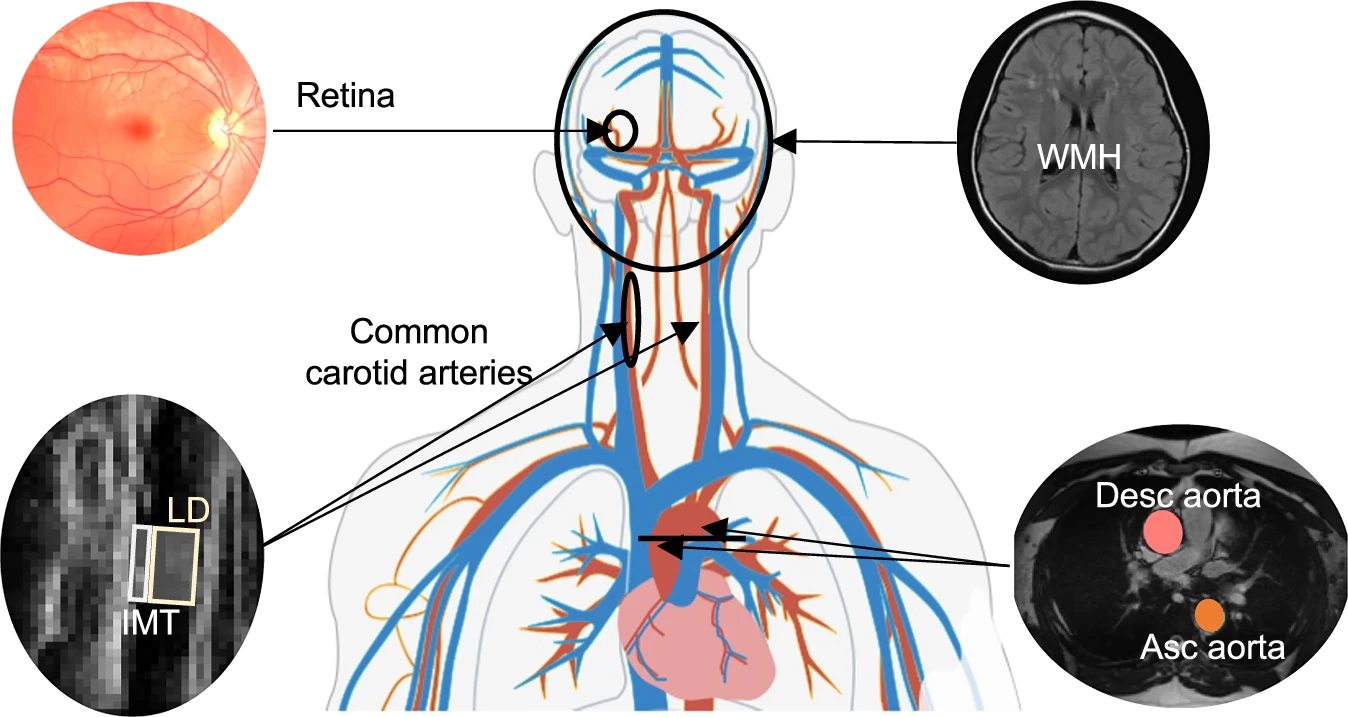
We must remember that vit.A , retina/brain/LRAT require phospholipids/lecithin
Lecithin forms cell mambranes and we lose it while aging
Perhaps some forms of vit.A "toxicity" could be a lecithin deficiency
AI
Lecithin supplementation has shown potential in supporting retinal function, primarily through its role in enhancing the bioavailability of nutrients essential for eye health and as a component of specialized medical treatments. Lecithin, particularly in the form of lecithin-bound iodine (LBI) or phosphatidylcholine, is associated with improvements in retinal pigment epithelium (RPE) health, tear film stability, and the reduction of oxidative damage in the retina.
Key Findings on Lecithin and Retinal Function:
Retinal Pigment Epithelium (RPE) Protection: Lecithin-bound iodine (LBI) has been used in Japan to treat retinal diseases, including central serous chorioretinopathy. In studies, LBI was shown to prevent hypoxic damage to RPE cells and suppress inflammatory responses (such as CCL2 secretion), suggesting a protective effect on the retinal structure.
Retinal Degeneration and Function: In animal models of retinal degeneration, a lecithin-containing diet improved retinal function, as evidenced by higher b-wave amplitude in electroretinography (ERG) tests compared to control groups. It was found to reduce the invasion of macrophages into the retina and decrease inflammatory markers.
Enhancing Nutrient Absorption: Lecithin acts as an emulsifier that helps the body absorb and utilize fat-soluble vitamins, including Vitamin A, which is crucial for producing visual pigments in the retina and maintaining photoreceptor function.
Exudative Age-Related Macular Degeneration (AMD): Research into the use of iodized lecithin tablets for exudative AMD indicated a potential positive impact, with studies reporting improved visual acuity and reduced retinal thickness in treated groups compared to control groups.
Tear Film Stability (Dry Eye Disease): Sunflower lecithin supplementation has been shown to improve dry eye disease (DED) symptoms. As a rich source of phosphatidylcholine, it may enhance the lipid secretions from the meibomian glands, stabilizing the tear film.
Retina Protection against Toxicity: Studies on rat retinas indicate that lecithin can help maintain the structural integrity of retinal proteins (specifically α-helix and β-sheet ratios) when exposed to oxidative stress, potentially reducing damage from, for example, chemotherapeutic agents like ifosfamide.
Important Context:
Lecithin-Retinol Acyltransferase (LRAT): Lecithin:retinol acyltransferase is an enzyme crucial for the metabolism of Vitamin A in the retina. It helps convert all-trans retinol to all-trans retinyl ester, a key step in the visual cycle.
Limitations: While promising, some uses of lecithin, specifically lecithin-bound iodine, have limited clinical trials outside of Japan, and many findings are based on preclinical or older studies.
In summary, lecithin supplementation, particularly when containing iodine or as a source of phosphatidylcholine, may support retinal function by improving structural integrity, reducing inflammation, and increasing the absorption of necessary nutrients, according to research.